
Oral glutathione supplementation drastically reduces Helicobacterinduced gastric pathologies
Ellen De Bruyne1, Richard Ducatelle1, Dennis Foss2, Margaret Sanchez2, Myrthe Joosten1, Guangzhi Zhang1, Annemieke Smet1, Frank Pasmans1, Freddy Haesebrouck1,* & Bram Flahou1,*
Helicobacter (H.) suis causes gastric pathologies in both pigs and humans. Very little is known on the metabolism of this bacterium and its impact on the host. In this study, we have revealed the importance of the glutamate-generating metabolism, as shown by a complete depletion of glutamine (Gln) in the medium during H. suis culture. Besides Gln, H. suis can also convert glutathione (GSH) to glutamate, and both reactions are catalyzed by the H. suis γ-glutamyltranspeptidase (GGT). Both for H. pylori and H. suis, it has been hypothesized that the degradation of Gln and GSH may lead to a deficiency for the host, possibly initiating or promoting several pathologies. Therefore the in vivo effect of oral supplementation with Gln and GSH was assessed. Oral supplementation with Gln was shown to temper H. suis induced gastritis and epithelial (hyper)proliferation in Mongolian gerbils. Astonishingly, supplementation of the feed with GSH, another GGT substrate, resulted in inflammation and epithelial proliferation levels returning to baseline levels of uninfected controls. This indicates that Gln and GSH supplementation may help reducing tissue damage caused by Helicobacter infection in both humans and pigs, highlighting their potential as a supportive therapy during and after Helicobacter eradication therapy.
Non-Helicobacter (H.) pylori Helicobacter (NHPH) species have been found colonizing the stomach of 0.2–6% of humans patients with severe gastric complaints1. Infection causes gastritis and peptic ulceration and the relative risk of developing mucosa-associated lymphoid tissue (MALT) lymphoma has been described to be higher with NHPH than with H. pylori1,2. Several studies have shown that H. suis, a member of the H. heilmannii sensu lato (s.l.) group3, is the most prevalent gastric NHPH in humans, which has been described to account for 14% to 78.5% of NHPH infections4–6. Interestingly, experimental infection studies in rodent models of human gastric disease have confirmed that long-term H. suis infection can lead to the development of gastric MALT lymphoma7–9. Besides humans, the majority of pigs worldwide are colonized by this bacterium, in which infection causes chronic gastritis and reduced average daily weight gain10. Transmission of H. suis most likely occurs through contact between pigs and humans1,11. Presence of viable H. suis bacteria in pork, however, suggests that foodborne infection might also occur12.
A persistent mild chronic gastritis can be observed in human patients after H. heilmannii s.l. eradication treatment with antibiotics and proton-pump-inhibitors11,13. For H. pylori, it has also been described that after successful eradication in humans, corpus gastritis and dysplasia improve, although they do not completely disappear. Especially antral lesions seem non-responsive after eradication therapy14. This emphasizes the need for supplements that promote health of the gastric mucosa.
Recently, we and others identified the γ-glutamyltranspeptidase (GGT) from H. suis and H. pylori as an important factor causing epithelial cell death and modulating lymphocyte responses15–18. For all gastric helicobacters, the mode of action of the GGT mostly depends on the breakdown of 2 substrates, glutathione (GSH) and glutamine (Gln), both leading to the production of glutamate8,19–22. Under certain conditions, GGT-mediated degradation of GSH facilitates the formation of reactive oxygen species, causing lipid peroxidation of cell membranes, which ultimately leads to apoptosis or necrosis of gastric epithelial cells in vitro15. Depletion of Gln in vitro can lead to an impaired proliferation and dysfunction of T-lymphocytes16–18.
Gln and GSH have a broad range of functions in the host. Gln plays a crucial role in the energy supply of rapidly dividing cells, such as intestinal epithelial cells and cells of the immune system23. In addition, this amino acid plays a role in protein turn-over and purine and pyrimidine synthesis24,25. The tripeptide GSH (γ-L-glutamyl-L-cysteinylglycine) is synthesized intracellularly. This ubiquitous free thiol is not only important for anti− oxidative protection of the plasma membrane and organelles, but it also plays distinct roles in cell cycle regulation and apoptosis26,27. After being transported outside the cells, GSH is degraded by eukaryotic membrane-bound GGT which removes the γ-glutamyl moiety28. The degradation products can be translocated into the cell, were they can for instance be recycled for GSH synthesis28.
Both deamination of Gln and degradation of GSH by Helicobacter GGT generate glutamate15,16,20, which can be taken up in the bacterial cell by a Na+-dependent transporter20,29. For H. pylori, metabolic reconstruction has suggested that glutamate can subsequently be converted to α -ketoglutarate, which can be channeled into the Tricarboxylic Acid cycle (TCA cycle)30. In contrast to most other bacteria, the TCA cycle of H. pylori is an atypical, non-cyclic pathway containing both an oxidative and reductive branch31. Some genes, necessary for a traditional TCA cycle are missing in the H. pylori genome. However, alternative reactions have been identified that allow connectivity between the metabolites of the TCA cycle30. No information is currently available on the metabolism of H. suis, a gastric Helicobacter species which is even far more fastidious than the well-studied H. pylori.
The present study is the first to examine the relative importance of the glutamate-generating metabolism (and by extension the amino acid metabolism) for survival and growth of H. suis. Since degradation of GSH and Gln by GGT from gastric helicobacters largely contribute to the glutamate-generating metabolism, we investigated the effect of GGT substrate supplementation (Gln/GSH) on the outcome of an experimental H. suis infection in Mongolian gerbils. By correcting for possible imbalances, we aimed at reducing infection-related gastric pathologies.
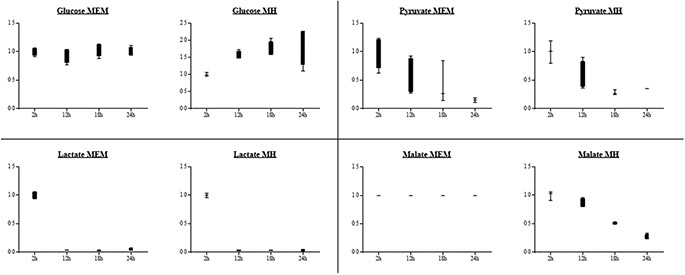
Figure 1. Carbohydrate concentrations in the supernatant of a liquid Helicobacter suis culture. Minimal Essential Medium (MEM) and Mueller Hinton (MH) medium. The y-axis represents the fold change of the different carbohydrates relative to the 2 hour measurement point.
Results
The glutamate-generating metabolism is vital for survival and growth of H. suis. Annotation of the H. suis genome identified no homologues for some important genes with a putative role in glucose metabolism21, suggesting that H. suis is unable to utilize the glycolytic pathway for metabolism. The genome of H. suis appears to lack glucose permease and several enzymes of the classic Embden-Meyerhof-Parnas pathway, including glucokinase and phosphofructokinase
21. Furthermore, most components of the Entner-Doudoroff pathway are absent in the H. suis genome, such as glucose 1-dehydrogenase and gluconate dehydratase
21. Utilization of glucose was not observed during H. suis culture in both Minimal Essential medium (MEM) and Mueller Hinton (MH) medium (Fig. 1, Table S1), further suggesting this bacterium does not utilize glucose as a major energy source. Instead, we hypothesized that this bacterium has to rely on other sources for its energy production. Pyruvate occupies a central branch point in the energy metabolism of H. suis. As the glycolytic pathway is impaired, pyruvate is most likely generated from amino acids such as serine, alanine and cysteine. This hypothesis was confirmed when we found that incubation of H. suis for 24 hours in MEM significantly lowered the extracellular concentrations of serine (p < 0.001), alanine (p < 0.001) and cysteine (p < 0.001). A decrease (up to 6-fold) of these amino acids was observed, probably due to the generation of pyruvate. In contrast, in MH-medium only cysteine concentrations significantly decreased (p < 0.001), whereas serine and alanine concentrations remained steady over time (Fig. 2). Genome annotation (using RAST) showed the presence of homologues for H. pylori genes encoding for L-serine deaminase (Accession number: NC.000915.1, gen ID 900301), cystathionine γ -synthase (NC.000915.1, gene ID 898970), alanine racemase (NC.000915.1, gen ID 899470) and D-amino acid dehydrogenase (NC.000915.1, gen ID 899472) in the H. suis genome (respective locus tags: HSUHS1_ RS00280, HSUHS1_RS06050, HSUHS1_RS00415 and HSUHS1_0061)21. Another amino acid that was decreased to basal levels after 24 h incubation in MEM was proline (p = 0.001, Fig. 2). We hypothesize that, as described for H. pylori, proline is taken up by the bacteria from the extracellular medium—mediated by the proline permease PutP32 – and subsequently converted to glutamate inside the bacteria by the proline utilizing flavoenzyme A (PutA)
32,33. A homologue of the putP gene of H. pylori (NC.00915.1, gen ID 899191) is indeed present in the genome of H. suis (locus tag HSUHS1_1153)21. No significant changes were detected for this amino acid when H. suis was incubated in MH-medium. Other substrates that decreased over time in the culture supernatant included malate and lactate. In MEM, lactate levels significantly decreased (p < 0.001). In MH-media, lactate, as well as malate concentrations decreased (resp. p = 0.001 and p < 0.001, Fig. 1). They showed up to a 7-fold reduction after H. suis culture, suggesting they may also be important carbon sources. A homologue for the H. pylori gene encoding for D-lactate dehydrogenase (NC.000915.1, gene ID 898825) is also present in the H. suis genome (locus tag HSUHS5_RS01825)21, allowing the conversion of lactate to pyruvate. Pyruvate can subsequently be converted to acetyl-CoA by pyruvate synthase. For H. pylori, it has been described that the main route for pyruvate assimilation is via a pyruvate:flavodoxin oxidoreductase (POR) (NC.00915.1, gen ID 899646)
34. ORF’s have been identified in the H. suis genome encoding a POR-type pyruvate synthase (pyruvate oxidoreductase: HSUSH1_RS06160, phosphoenolpyruvate synthase: HSUHS1_RS07385)21, suggesting that a similar mechanism is present in H. suis. As described for H. bizzozeronii, the genome of H. suis also codes for phosphoenol pyruvate carboxylase (locus tag HSUHS1_RS00640)21, an enzyme that plays a role in the synthesis of oxaloacetate from phosphoenolpyruvate35. Certain amino acids were nearly depleted from the medium of a 24 h culture of H. suis. The levels of asparagine decreased approximately 50-fold (p < 0.001, Fig. 2), while the level of Gln dropped below the detection limit (p < 0.001, Fig. 2). Metabolic reconstruction showed that homologs for HP-ansB/HP-dcuA and HP-ggt/ HP-gltS are present in the H. suis genome21. For H. pylori, these hydrolysis/transport systems have been shown to be responsible for the deamination of asparagine and Gln and subsequent uptake by the bacteria of aspartate and glutamate, respectively
29. These amino acids not only serve as precursors to protein synthesis, they can also be metabolized as energy source via the TCA cycle. Indeed, a homologue for the H. pylori gene coding for NADP-dependent glutamate dehydrogenase (NC.000915.1, gen ID 898871) is present in the genome of H. suis (locus tag HSUHS1_RS07485)
21. This enzyme can convert glutamate to 2-ketoglutarate, an intermediate metabolite of the TCA cycle, whereas aspartate can be channeled into the TCA cycle by aspartate ammonia-lyase or other multi-step pathways, such as adenylosuccinate synthase—adenylosuccinate lyase. In any case, this underlines the importance of the amino acid metabolism and Gln metabolism in particular for the growth and energy supply of H. suis. A more detailed report of the metabolites used by H. suis in both the MEM- and MH-medium can be found in supplementary Table S1.
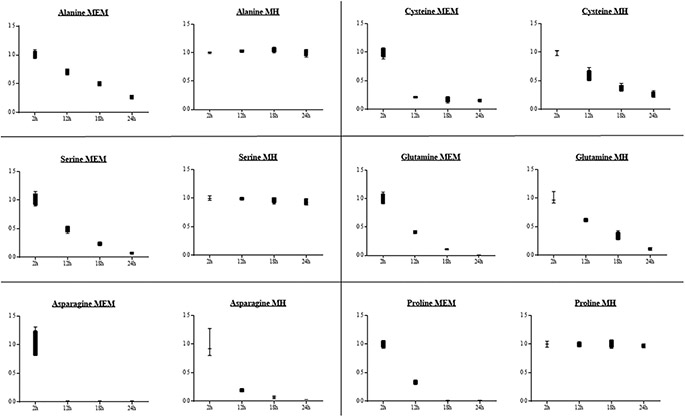
Figure 2. Amino acid concentrations in the supernatant of a liquid Helicobacter suis culture. Shown are the relative fold changes of the different amino acid concentrations compared to the 2 hour measurement point. MEM = Minimal Essential Medium, MH = Mueller Hinton.
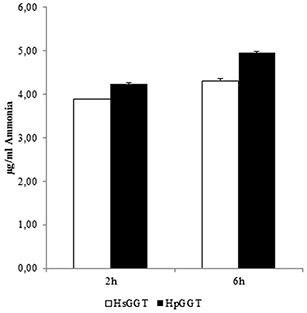
Figure 3. Production of ammonia during Helicobacter suis/H. pylori GGT-mediated degradation of glutamine.
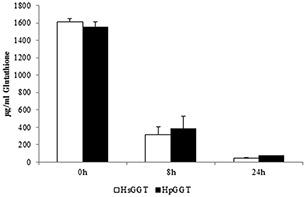
Figure 4. Degradation of GSH by Helicobacter suis GGT (HsGGT) and H. pylori GGT (HpGGT). After supplementing 2 μ g/ml HsGGT or HpGGT to Ham’s F12 containing 1600 µg/ml glutathione (GSH), a decrease in GSH concentration was observed.
Degradation of Gln and GSH by Helicobacter suis GGT
H. suis GGT (HsGGT) and H. pylori GGT (HpGGT) were shown to have a very similar Gln- and GSH-degrading capacity. After 2 hours of incubation, 3.88 ± 0.01 μ g/ml and 4.23 ± 0.02 μ g/ml ammonia was generated by Gln degradation through 2 μ g/ml HsGGT and HpGGT, respectively. After 6 hours of incubation, slightly higher amounts of ammonia were generated (4.30 ± 0.06 μ g/ml and 4.95 ± 0.02 μ g/ml, respectively) (Fig. 3). After 8 hours of incubation, the initial GSH concentration was reduced by 80%, which added up to 97% after 24 h. Comparable data were obtained for HpGGT. After 8 h, the GSH concentration was reduced by 75%, whereas after 24 h the GSH concentration was reduced by 95% (Fig. 4).
GSH and Gln supplementation drastically reduces Helicobacter-induced inflammation and cell damage. Given the importance of the glutamate-generating metabolism for H. suis and the potential impact of the concurrent Gln and GSH degradation (with an important role for H. suis GGT in these processes), we investigated the effects of feed supplementation with both substrates to H. suis infected Mongolian gerbils. No differences in feed intake were observed between animals from different groups during the experiment (p > 0.05, Table S2). In addition, animals in different groups showed a similar weight gain during the course of the experiment (Table S2). All uninfected control animals tested negative for the presence of H. suis. In experimentally infected animals, the highest H. suis colonization rates were found in the antrum. Average colonization rates in animals receiving the standard diet, Gln-supplemented and GSH-supplemented diet were 2.49 × 104 (± 2.76 × 104), 1.13 × 104 (± 6.08 × 104) and 2.62 × 103 (± 5.53 × 103) bacteria/mg tissue, respectively. There were no statistically significant differences between different groups, although animals receiving the GSH diet tended to show lower colonization rates (Standard vs Gln p = 0.606; Standard vs GSH p = 0.129, Fig. S1).
None of the control animals showed signs of gastritis. In H. suis infected animals, gastritis was mainly present in the antrum. There was a significant difference in gastritis scores between the different groups of infected animals (Fig. 5). A detailed overview of the observed inflammation scores can be found in the online supplementary data (Table S3). Infected animals belonging to the group receiving the GSH supplemented diet showed substantially lower inflammation scores (mean score 0.3 ± 0.4; Fig. 6D) compared to infected animals from the group receiving the purified standard diet (mean score 2.6 ± 0.4; p < 0.001; Fig. 6B). Seven out of 10 H. suis infected animals receiving the GSH-supplemented diet even showed no detectable inflammation. Although notably less lymphocytic infiltration and fewer lymphocytic aggregates were also observed in the infected animals receiving the Gln supplemented diet (mean score 1.9 ± 0.9; Fig. 6C), this was not statistically significant (p = 0.064). In general, deep lymphocytic infiltrates consisted mainly of CD3-positive cells whereas superficial lymphoid follicles contained a majority of CD3-negative B lymphocytes (Fig. S2). Results of cytokine expression are summarized in Fig. 7. For most cytokines tested, no differences were observed between control groups receiving different diets. Surprisingly, however, uninfected control animals receiving the Gln-supplemented diet did show an upregulation of INF-γ expression (p = 0.012, fold = 3.01 ± 1.19). When comparing control and H. suis infected animals, both receiving the standard diet, H. suis infection was shown to provoke an upregulation of IL-1β , INF-γ and IL-10 expression. In H. suis infected animals receiving the Gln-supplemented diet, only an upregulation of IL-1β was noted, whereas mRNA expression levels of other cytokines dropped back to the levels of uninfected control animals.
Animals receiving the GSH diet even showed cytokine expression levels comparable to those in uninfected control animals. No differences in epithelial cell proliferation were observed between the non-infected groups receiving different diets. Marked differences in terms of gastric epithelial proliferation at the level of the antral pits were observed between H. suis infected groups receiving different diets (Figs 8 and 9). H. suis infected animals receiving the standard diet showed the highest epithelial proliferation rate (8.2 ± 1.0 Ki67-positive cells per gland; p < 0.001). The proliferation rate was significantly lower in animals receiving the Gln- (6.2 ± 1.1, p = 0.010) or the GSH-supplemented diet (4.9 ± 1.6, p = 0.002) compared to infected animals receiving the non-supplemented diet. In the corpus, no differences in epithelial cell proliferation were observed between the different H. suis infected groups (Fig. 9). Besides their abundant presence in the corpus, a limited number of parietal cells can also be found in the antrum. An abrupt loss of parietal cells was observed in the transition zone between the corpus and antrum in the H. suis infected animals receiving the standard diet as well as the Gln-supplemented diet. In H. suis infected animals receiving the GSH-supplemented diet, this abrupt loss was absent, revealing an image comparable to that of uninfected control animals (Fig. S3).
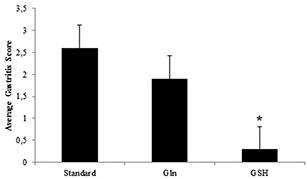
Figure 5. Average gastritis scores observed in the antrum of the stomach of H. suis infected animals shows a markedly decreased inflammation in the group receiving the glutathione diet (GSH). An *depicts significantly lower values compared to the H. suis infected animals receiving the standard feed. Error bars represent the standard deviation.
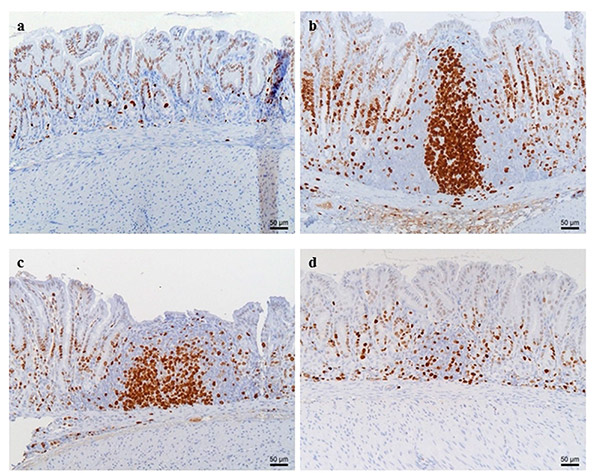
Figure 6. Proliferating lymphocytes in the antrum of control (a)and Helicobacter suis infected animals (b–d). Proliferation of lymphocytes, as well as the formation of lymphoid follicles with proliferating germinal centers, is observed in H. suis infected animals receiving the standard diet (b) and infected animals receiving the glutamine-supplemented diet (c). In contrast, only a limited amount of proliferating lymphocytes are observed in H. suis infected animals receiving the diet supplemented with glutathione (d). In control animals (a), only a limited number of cells are proliferating and no lymphocytic aggregates are present. Proliferating Ki-67 positive cells are brown.
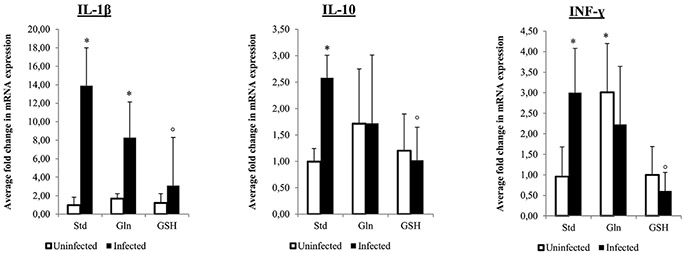
Figure 7. Fold change in cytokine expression in control and Helicobacter suis infected animals. An *indicates a significant difference in infected animals compared to the negative control animals receiving the standard diet. An °indicates a significant difference in expression compared to H. suis infected animals receiving the standard diet (p < 0.05). Std: animals receiving the standard diet; Gln: Glutamine-supplemented diet; GSH: Glutathionesupplemented diet.
Discussion
Metabolic profiling and the absence of putative genes encoding important enzymes of the Embden-Meyerhof- Parnas and Entner-Doudoroff pathway suggest that, in contrast to the closely related H. pylori and H. bizzozeronii, H. suis is not able to utilize glucose for its main energy supply. As a consequence, H. suis has to rely on other sources to fulfill its energy requirements. Metabolic reconstruction and analysis of metabolites in H. suis culture medium showed that H. suis tends to use amino acids, not only for protein synthesis, but most likely also as an energy source. The consumption of amino acids was obvious in cultures grown in both liquid media, but was most pronounced in MEM, which can be considered to be the most basic one of the two. This could indicate that H. suis relies even more on amino acids for its survival when facing stressful situations. One of the preferred nutrients seems Gln, which is the most abundant free amino acid in both plants and animals36–38. Our current and previous studies have shown that Gln is depleted from the medium during H. suis culture due to the hydrolysis by H. suis GGT16. As described for H. pylori, the formed glutamate can be taken up and converted to 2-ketoglutarate by the bacteria and channeled into the TCA cycle21,29, an important metabolic pathway that provides precursors for a variety of cell components, as well as for supplying energy to the bacterium31.
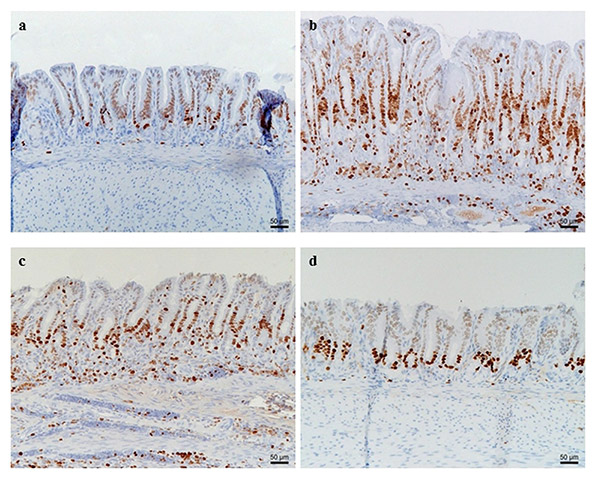
Figure 8. Proliferating Ki67-positive (brown) epithelial cells and lymphocytes in the antrum of the stomach of control and Helicobacter suis infected animals. (a) Control animal receiving the standard diet; (b) H. suis infected animal receiving the standard diet; (c) H. suis infected animal receiving the glutaminesupplemented diet; (d) H. suis infected animal on the glutathione-supplemented diet. A more pronounced proliferation is observed in H. suis infected animals receiving the standard and glutamine-supplemented diet compared to H. suis infected animals receiving the glutathione-supplemented diet.
The hydrolysis of extracellular Gln by the GGT from gastric helicobacters can cause a depletion of this conditionally essential amino acid in the gastric juice of an infected host, which may eventually lead to epithelial cell damage and immunological dysfunction16,18,20. By supplementing Gln to the diet of Mongolian gerbils we aimed at reducing inflammation-related as well as other Helicobacter-related gastric pathological changes. In addition to a local effect, dietary Gln supplementation has been described to cause an increase of the systemic Gln concentration, which may also sustain the health and function of immune and epithelial cells39. The role of Gln in gastric mucosal protection against H. pylori infection has been reported previously36–38,40–42. In these studies, an effect of Gln supplementation on gastric inflammation and cytokine expression was observed41. In addition, a recent study indicated that the depletion of Gln, as a result of high GGT activity, might actually be responsible for the strong inflammatory response following H. pylori infection, as shown by increased release of pro-inflammatory IL-8 from epithelial cells17. In contrast, Wüstner et al.18 described that H. pylori GGT activity leads to an impaired T-cell function by depriving activated lymphocytes from Gln. In the present study, the beneficial effect of Gln supplementation on inflammation was only mild to moderate, which may be due to the shorter infection period compared to that described by Hagen et al.41. Nevertheless, H. suis infected animals receiving the Gln diet showed smaller gastric lymphoid follicles and less diffuse lymphoid infiltration compared to the animals receiving the standard diet. This decreased proliferation of lymphocyte populations, as well as the reduced expression of pro- and anti-inflammatory cytokines in H. suis infected animals receiving Gln-supplemented feed is somewhat in contrast to in vitro studies showing that Gln supplementation restores the impaired proliferation of H. suis GGT-treated Jurkat T-cells16.
Other factors are most likely involved, such as a protective effect of Gln on the epithelial barrier. Indeed, the current study is the first to show a direct effect of Gln supplementation on the proliferation rate of gastric epithelial cells in the context of a Helicobacter infection, suggesting a protective effect of Gln on gastro-intestinal epithelial cells43–45. Most likely, hyperproliferation of epithelial cells found in H. suis infected animals receiving the standard diet is a secondary response to increased cell death induced by this bacterium8,45,46. Increased cell death may be induced by an increased oxidative stress burden or a modification of the redox balance of gastric epithelial cells following Gln depletion15,47. Supplementing Gln may therefore assist in restoring the redox balance. Besides oxidative stress, H. suis infection causes a dramatic increase of ammonia concentrations, resulting from urease and GGT activity of the bacterium1,20,42,48. Adding Gln to the diet can stimulate arginase activity, causing a conversion of ammonia to the less toxic urea, and helping in clearing ammonia from the cells by stimulating its excretion23,42. Although it apparently has a beneficial effect on compensatory hyperproliferation of gastric epithelial cells, supplementing Gln to H. suis infected animals does not prevent the abrupt loss of parietal cells at the transition zone between the corpus and antrum8. Besides a depletion of Gln, GGT from gastric helicobacters also causes degradation of GSH15,16. Previous in vitro research has revealed that supplementation of GSH at a concentration of 5 mM to H. suis GGT-treated epithelial cells can stimulate the formation of pro-oxidative GSH degradation products and extracellular oxygen radicals, leading to necrosis of damaged cells15. In the present study, however, supplementing 0.8% GSH to the diet of H. suis infected gerbils revealed tremendously beneficial effects on the health of a Helicobacter-infected stomach.
We observed a marked decrease of gastric epithelial cell proliferation rates, protection against parietal cell death and a 90% reduction of gastritis, accompanied by normalized expression levels for all (pro-inflammatory) cytokines, confirming the suppression of the host immune response elicited by Helicobacter infection. Only three animals had small lymphoid follicles in the antrum of the stomach, despite normal H. suis colonization rates. Helicobacter GGT has been shown to inhibit proliferation of T-lymphocytes, an effect that has been suggested to play a role in the immune escape by these bacteria16,49. Supplementing GSH to the cell medium has been shown to aggravate this effect16. Possibly, similar mechanisms are involved in the current in vivo study. The observed protective effects on infection-induced inflammation and, surprisingly, epithelial cell hyperproliferation may be explained by a dose-dependent effect of GSH. Intracellular concentrations of GSH are usually very high, ranging from 0.5 to 10 mM, whereas extracellular concentrations are found in the micromolar range50. Therefore, supplementing feed with higher concentration of GSH (35 mM) compared to the dosages used for previous in vitro studies (5 mM)15 probably results in an excess quantity of protective extracellular GSH, most likely counterbalancing some potentially negative effects on gastric epithelial cell health by GGT-induced generation of pro-oxidative GSH degradation products15. It has indeed been shown that, when available for the host cells, GSH plays an important role in the protection of cells against oxygen radicals, detoxification and metabolisation of various endo- and exogenous compounds, cell cycle regulation, cell signaling and apoptosis16,50–52. GSH deficiency makes cells more sensitive to oxidative stress and can cause degeneration of epithelial cells50,51. The remarkable beneficial effects of GSH on Helicobacter-induced inflammation and epithelial cell homeostasis suggest that GSH supplementation could be considered as a supportive therapy during and even after a Helicobacter eradication therapy.
It should be noted, however, that only the short/medium-term effects of GSH supplementation were evaluated during this trial. A study of Liu et al.53 showed that GSH supplementation had beneficial effects on acute clinical signs of inflammatory bowel disease (IBD), whereas some adverse effects were observed in the chronic stages of IBD in this particular disease model. More research is therefore needed to evaluate the long-term safety and effects of GSH supplementation and to make recommendations on the chronic use of glutathione in people suffering from gastric disease. Feed additives are commonly used in the pig industry to promote pig health. They are easily manufactured and provided to the animals. Both Gln and GSH can be considered as a possible feed additive since the concentrations of both were shown to be stable in the feed pellets used in the present study (unpublished results). These supplements may therefore be helpful for decreasing both the prevalence and severity of gastric pathologies caused by H. suis infection in pigs. Indeed, up to 90% of H. suis infected pigs have been shown to develop gastritis, which most likely plays a role in the observed reduction of daily weight gain, a primordial production characteristic in the pig industry10. In conclusion, we showed that H. suis preferably uses amino acids to meet its energy requirements. In particular, the glutamate-generating metabolism seems essential, as shown by a complete depletion of Gln during H. suis culture. The H. suis GGT has previously been shown to be an important factor degrading Gln, but also GSH. Oral supplementation of H. suis infected animals with these two substrates, Gln or GSH, resulted in a marked reduction of infection-related gastric pathologies, both related to inflammation and epithelial cell damage. GSH was shown to be superior to glutamine and may prove to be of therapeutic value for use in both humans and animals. Methods Metabolic Profiling of H. suis: Energy Metabolism. H. suis type strain HS1LP was grown in liquid Minimal Essential Medium (MEM) and Mueller Hinton (MH) broth.
The sample preparation process was carried out using the automated MicroLab STAR® system (Hamilton Company, Reno, US). Recovery standards were added prior to the first step in the extraction process for Quality Control purposes. Sample preparation was conducted using a proprietary (Metabolon, Durham, North Carolina, USA) series of organic and aqueous extractions to remove the protein fraction while allowing maximum recovery of small molecules. The resulting extract was divided into two fractions, one for analysis by Liquid Chromatography (LC) and one for analysis by gas chromatography (GC). Samples were placed briefly on a TuboVap® (Zymark, Massachusetts, US) to remove the organic solvent. Each sample was then frozen and dried under vacuum. Samples were then prepared for the appropriate instrument, either LC/MS or GC/MS. The LC/MS portion of the platform was based on a Surveyor HPLC and a Thermo-Finnigan LTQ Fmass spectrometer (Thermo scientific, Waltham, Massachusettes, USA), which consisted of an electrospray ionization (ESI) source and linear ion-trap (LIT) mass analyzer. Positive and negative ions were monitored within a single analysis by consecutively alternating the ionization polarity of adjacent scans. The vacuum-dried sample was dissolved in 100.0 μ l of an injection solvent that contained five or more injection standards at fixed concentrations. The chromatography has been standardized by Metabolon, Inc. and once a method was validated, no changes were made to it. The internal standards were used both to assure injection and chromatographic consistency. The chromatographic system used a binary solvent (water and methanol) system delivered as a gradient. The HPLC columns were washed and reconditioned after every injection. The samples destined for GC/MS analysis were re-dried under vacuum desiccation for a minimum of 24 hours prior to being derivatized under dried nitrogen using bistrimethyl-silyl-triflouroacetamide (BSTFA). The GC column was 5% phenyl and the temperature ramp ranged from 40° to 300 °C in a 16 minute period. Samples were analyzed on a Thermo-Finnigan Trace DSQ fast-scanning single-quadrupole mass spectrometer using electron impact ionization. At 2, 12, 18 and 24 hours media supernatants was analyzed. For each condition, 4 biological replicates were performed. A small aliquot of each experimental sample for a specific matrix was obtained and pooled. These pooled samples were injected throughout the platform day run and served as technical replicates. In total, 702 media were identified and statistical analysis was performed to detect concentration changes during H. suis culture. Based on information derived from a draft genomic sequence21, a partial metabolic reconstruction of H. suis strains HS1 and HS5 (currently named H. suis strains HS1LP and HS5bLP54) was performed, mainly focusing on the carbon metabolism of this bacterium and attempting to confirm the results of metabolic profiling (Integrated Genomics Inc., Chicago, Illinois, US). In addition, the RAST (Rapid Annotation using Subsystem Technology) tool was used to confirm the presence/absence of genes important for the metabolism of H. suis in the genome of H. suis strains HS1LP and HS5bLP21,54.
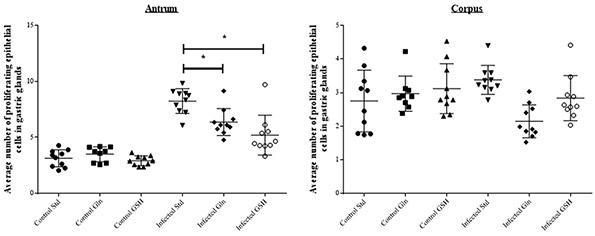
Figure 9. Proliferation of gastric epithelial cells. Ki67 staining of the stomach epithelium in the antrum and corpus of the stomach. Std: standard feed, Gln: glutamine-supplemented feed, GSH: glutathione-supplemented feed. An *depicts significantly lower values compared to the H. suis infected animals receiving the standard feed. Error bars represent the standard deviation.
Effects of H. suis and H. pylori GGT on Gln and GSH
The H. suis and H. pylori GGT were expressed and purified as described previously15. Both enzymes were compared to each other regarding their capacity to degrade 2 important substrates: Gln and GSH. Two μ g/ml of H. suis or H. pylori GGT was added to HBSS supplemented with 2 mM Gln and incubated at 37 °C for 2 hours and 6 hours. As a control, a solution of HBSS supplemented with 2 mM Gln was used. The concentration of produced ammonia was detected using an Ammonia Assay Kit (Abcam, Cambridge, UK) according to the manufacturer’s instructions. To evaluate the effect of HsGGT and HpGGT on GSH, 1600 µg/ml GSH was incubated for 8–24 hours at 37 °C with 2 μ g/ml HsGGT or HpGGT in Ham’s F12 medium. GSH concentration was measured using the Glutathione Assay Kit (Sigma-Aldrich) according to the manufacturer’s instructions.
Bacterial strain used for in vivo experimentation.
H. suis strain HS5bLP, isolated from the gastric mucosa of a sow55, was used for experimental infection of Mongolian gerbils. Culture was performed as described previously54.
Ethics statement
All experimental procedures were approved by the Ethical Committee of the Faculty of Veterinary Medicine of the University of Ghent, Belgium (approval number EC2011-113), and carried out in accordance with the approved guidelines and regulations.
Animals, housing, inoculation and sampling
Sixty female Mongolian gerbils (Charles River, France) arrived at our animal housing units at the age of 4 weeks. They were randomly divided in 6 groups with 10 animals each. Animals from 1 group were divided in 2 filter-top cages (5 animals each), all held in the same stable under controlled environmental conditions. During the experiment, feed intake was monitored weekly and body weight was measured three times, twice at inoculation and once at euthanasia. Three different rodent diets (Table 1) were provided ad libitum to 20 animals each (Research Diets, Inc., New Brunswick, USA). The standard diet was a purified feed (AIN-93 G, Research Diets, Inc.). The other 2 diets were identical to the standard diet, with the exception of being supplemented with 5% Gln or 0.8% GSH and with a variable amount of corn starch, used to correct minor differences in the energy content. At the age of 6 weeks, the animals were briefly anaesthetized with 3% isoflurane and inoculated intragastrically twice, with a two-day interval. Ten animals from each feed group received 8 × 107 viable H. suis bacteria/dose. The remaining 10 animals/feed group were sham-inoculated with the culture medium of H. suis. Twelve weeks after the first inoculation, animals were anaesthetized with 5% (vol/vol) isoflurane followed by cervical dislocation. The stomach was opened along the greater curvature. A longitudinal strip was taken from the forestomach to the duodenum, fixed in 4% phosphate buffered formaldehyde and embedded in paraffin for histopathological examination. Biopsies from antrum and corpus were taken for DNA and RNA extraction. These tissues were immediately submerged in RNAlater (Qiagen, Hilden, Germany) and stored at −70 °C until further processing. DNA extraction and quantitative-PCR to determine the colonization rate of H. suis. DNA extraction and Quantitative-PCR were performed on the DNA samples as described previously56,57.
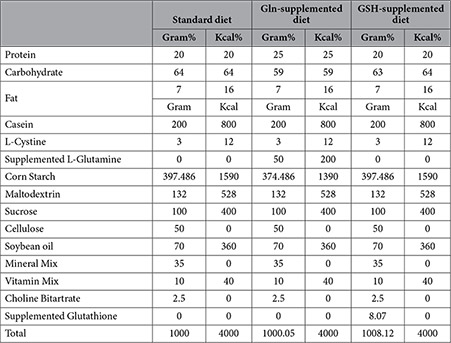
Table 1. Composition of the 3 different diets used in this study.
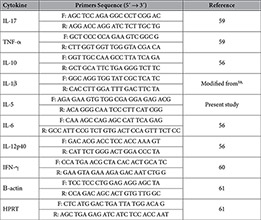
Table 2. Forward (F) and reverse (R) primers used for cytokine-expression.
RNA extraction and Real-Time PCR analysis of cytokine expression
After separation of DNA and RNA, the latter was further purified using the RNeasy Mini kit (Qiagen) following the suppliers’ instructions. Synthesis of cDNA was done by using the iScriptTM cDNA synthesis kit (Bio-rad, Hercules, USA) as described by the manufacturer. Real Time− PCR and melting curve analysis were done as described before58. The primer sequences are summarized in Table 2. Primers for interleukin (IL)-5 were designed using primer3 software.
Histopathology
Four sections of 5 μ m were sliced from the paraffin-embedded stomach tissue. The first section was used for a hematoxylin and eosin (HE) staining to score the intensity of infiltration with mononuclear and polymorphonuclear cells. A second tissue slide was stained using a mouse monoclonal anti-Ki67 antibody (Menarini Diagnostics, Firenze, Italy; dilution 1:50) and a biotinylated goat anti-mouse IgG antibody to identify proliferating cells. Positive cells lining the lumen of the gastric pits were counted in five randomly chosen High Power Fields (magnification: x400) in the corpus and antrum. For each animal, an average of the positive cell count was determined for both stomach regions. A third section was used to stain acid-producing parietal cells of the corpus of the stomach by using an antibody directed against the H+K+ATP− ase β -subunit (Abcam, Cambridge, UK; dilution 1:25000). The fourth tissue slide was used for visualization of T-lymphocytes, by using an antibody directed against CD3 (Abcam, dilution 1:100).
Statistical analysis
T-tests were used to analyze the data of the metabolomics study. T-test comparisons were performed between each experimental group and its control. Differences in gastritis between experimentally infected animals and control animals receiving the standard diet were analyzed using a Mann-Whitney U non-parametric test. Comparison of the colonization rates, cytokine expression and epithelial proliferation rates between the different diet groups of infected animals was done by means of a one-way ANOVA (SPSS 21, IBM, New York, USA). P-values ≤ 0.05 were considered statistically significant. When multiple groups were compared, a Bonferroni correction was made. All data are expressed as mean ± standard deviation.
References
1. Haesebrouck, F. et al. Gastric helicobacters in domestic animals and nonhuman primates and their significance for human health. Clin. Microbiol. Rev. 22, 202–223 (2009).
2. Stolte, M. et al. Helicobacter and gastric MALT lymphoma. Gut 50, suppl. iii19–iii23 (2002).
3. Haesebrouck, F. et al. Non-Helicobacter pylori Helicobacter species in the human gastric mucosa: a proposal to introduce the terms H. heilmannii sensu lato and sensu stricto. Helicobacter 16, 339–340 (2011).
4. Trebesius, K., Adler, K., Vieth, M., Stolte, M. & Haas, R. Specific detection and prevalence of Helicobacter heilmannii-like organisms in the human gastric mucosa by fluorescent in situ hybridization and partial 16S ribosomal DNA sequencing. J. Clin. Microbiol. 39, 1510–1516 (2001).
5. De Groote, D. et al. Detection of non-Helicobacter pylori species in “Helicobacter heilmanni”-infected humans. Helicobacter 10, 398–406 (2005).
6. Van den Bulck, K. et al. Identification of non-Helicobacter pylori spiral organisms in gastric samples from humans, dogs, and cats.
J. Clin. Microbiol. 43, 2256–2260 (2005).
7. Nakamura, M. et al. “Candidatus Helicobacter heilmannii” from a cynomolgus monkey induces gastric mucosa-associated lymphoid tissue lymphomas in C57BL/6 mice. Infect. Immun. 75, 1214–1222 (2007).
8. Flahou, B. et al. Helicobacter suis causes severe gastric pathology in mouse and mongolian gerbil models of human gastric disease. PLoS One 5, e14083 (2010).
9. Nakamura, M. et al. Microcirculatory alteration in low-grade gastric mucosa-associated lymphoma by Helicobacter heilmannii infection: Its relation to vascular endothelial growth factor and cyclooxygenase-2. J. Gastroenterol. Hepatol. 23, S157–S160 (2008).
10. De Bruyne, E. et al. An experimental Helicobacter suis infection causes gastritis and reduced daily weight gain in pigs. Vet. Microbiol. 160, 449–454 (2012).
11. Joosten, M. et al. Case report: Helicobacter suis infection in a pig veterinarian. Helicobacter 18, 392–396 (2013).
12. De Cooman, L. M. et al. Survival of Helicobacter suis bacteria in retail pig meat. Int. J. Food Microbiol. 1, 164–167 (2013).
13. Morgner, A. et al. Helicobacter heilmannii-associated primary low-grade MALT lymphoma: complete remission after curing the infection. Gastroenterol 118, 821–828 (2000).
14. Kokkola, A. et al. The effect of Helicobacter pylori eradication on the natural course of atrophic gastritis with dysplasia. Aliment. Pharmacol. Ther. 16, 515–520 (2002).
15. Flahou, B. et al. Gastric epithelial cell death caused by Helicobacter suis and Helicobacter pylori gamma-glutamyl transpeptidase is mainly glutathione degradation-dependent. Cell. Microbiol. 13, 1933–1955 (2011).
16. Zhang, G. et al. Effects of Helicobacter suis γ - glutamyl transpeptidase on lymphocytes: modulation by glutamine and glutathione supplementation and outer membrane vesicles as a putative delivery route of the enzyme. PLoS One 10, e77966 (2013).
17. Rimbara, E., Mori, S., Kim, H. & Shibayama, K. Role of γ-glutamyltranspeptidase in the pathogenesis of Helicobacter pylori infection. Microbiol. Immunol. 57, 665–673 (2013).
18. Wüstner, S. et al. Helicobacter pylori γ-glutamyltranspeptidase impairs T-lymphocyte function by compromising metabolic adaption through inhibition of cMyc and IRF 4 expression. Cell. Microbiol. 17, 51–61 (2015).
19. Chevalier, C., Thiberge, J. M., Ferrero, R. L. & Labigne, A. Essential role of Helicobacter pylori γ-glutamyltranspeptidase for the colonization of the gastric mucosa of mice. Mol. Microbiol. 31, 1359–1372 (1999).
20. Shibayama, K. et al. Metabolism of glutamine and glutathione via γ-glutamyltranspeptidase and glutamate transport in Helicobacter pylori: possible significance in the pathophysiology of the organism. Mol. Microbiol. 64, 396–406 (2007).
21. Vermoote, M. et al. Genome sequence of Helicobacter suis supports its role in gastric pathology. Vet. Res. 42, 51 (2011).
22. Rossi, M. et al. Evidence of conserved function of γ-glutamyltranspeptidase in Helicobacter Genus. PloS One 7, e30543 (2012).
23. Aledo, C. J. Glutamine breakdown in rapidly dividing cells: waste or investment? Bioessays 26, 778–785 (2004).
24. DeBerardinis, R. J. & Cheng, T. Q’s next: the diverse functions of glutamine in metabolism, cell biology and cancer. Oncogene 29, 313–324 (2010).
25. Hsu, C. B., Huang, H. J., Wang, C. H., Yen, W. H. T. & Yu, B. The effect of glutamine supplement on small intestinal morphology and xylose absorptive ability of weaned piglets. Afr. J. Biotechnol. 9, 7003–7008 (2010).
26. Shibayama, K. et al. Apoptotic signaling pathway activated by Helicobacter pylori infection and increase of apoptosis-inducing activity under serum-starved conditions. Infect. Immun. 69, 3181–3189 (2001).
27. Zhang, H. & Forman, H. J. Redox regulation of gamma-glutamyl transpeptidase. Am. J. Resp. Cell. Mol. 41, 510–515 (2009).
28. Drozdz, R. et al. Gamma-glutamyltransferase dependent generation of reactive oxygen species from a glutathione/transferrin system. Free Radical Bio. Med. 25, 786–792 (1998).
29. Leduc, D., Gallaud, J., Stingl, K. & De Reuse, H. Coupled amino acid deamidase-transport systems essential for Helicobacter pylori colonization. Infect. Immun. 78, 2782–792 (2010).
30. Schilling, C. H. et al. Genome-scale metabolic model of Helicobacter pylori 26695. J. Bacteriol. 184, 4582–4593 (2002).
31. Pitson, S. M., Mendz, G. L., Srinivasan, S. & Hazell, S. L. The tricarboxylic acid cycle of Helicobacter pylori. Eur. J. Biochem. 1, 258–267 (1999).
32. Krishnan, N. & Becker, D. F. Oxygen reactivity of PutA from Helicobacter species and proline-linked oxidative species. J. Bacteriol. 188, 1227–1235 (2006).
33. Nagata, K. et al. L-serine, D- and L-proline and alanine as respiratory substrates of Helicobacter pylori: correlation between in vitro and in vivo amino acid levels. Microbiology 149, 2023–2030 (2003).
34. Hughes, N. J., Clayton, C. L., Chalk, P. A. & Kelly, D. J. Helicobacter pylori porCDAB and oorDABC genes encode distinct pyruvate:flavodoxin and 2-oxoglutarate: acceptor oxidoreductases which mediate electron transport to NADP. J. Bacteriol. 180, 1119–1128 (1998).
35. Scott, T., Kondadi, P. K., Hänninen, M. & Rossi, M. Comparative Genomics of Helicobacter pylori and the human-derived Helicobacter bizzozeronii CIII-1 nature of non-pylori gastric Helicobacter infections in humans. BMC Genomics 12, 534 (2011). 36. Lam, H. M., Coschigano, K. T., Oliveira, I. C., MeloOliveira, R. & Coruzzi, G. M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 569–593 (1996). 37. Roth, E., Spittler, A. & Oehler, R. Glutamine: Effects on the immune system, protein metabolism and intestinal function. Wien. Klein. Wochensch. 108, 669–676 (1996).
38. Wu, G. et al. Important roles for L-glutamine in swine nutrition and production. J. Anim. Sci. 89, 2017–2030 (2011). 39. Vicarcio, M. et al. Dietary glutamine affects mucosal functions in rats with mild DSS-induced colitis. J. Nutrition. 137, 1931–1937 (2007). 40. Nakamura, E. et al. Prophylactic effect of glutamine and marzulene against Helicobacter pylori-related gastric diseases in Mongolian gerbils. Jpn. Pharmacol. Ther. 35, 479–488 (2007). 41. Hagen, S. et al. Inflammation and foveolar hyperplasia are reduced by supplemental dietary glutamine during Helicobacter pylori infection in mice. J. Nutr. 22, 912–918 (2009). 42. Amagase, A. et al. New frontiers in gut nutrient sensor research: prophylactic effect of glutamine against Helicobacter pylori-induced gastric diseases in Mongolian gerbils. J. Pharmacol. Sci. 112, 25–32 (2010). 43. Nakamura, E. & Hagen, S. Role of glutamine and arginase in protection against ammonia-induced cell death in gastric epithelial cells. Am. J. Physiol-Gastr. L. 283, G1264–G1275 (2002). 44. Evans, M. E., Jones, D. P. & Ziegler, T. R. Glutamine prevents cytokine-induced apoptosis in human colonic epithelial cells. J. Nutr. 133, 3065–3073 (2003).
45. Wagner, S. et al. Regulation of gastric epithelial cell growth by Helicobacter pylori: Evidence for a major role of apoptosis. Gastroenterol. 113, 1836–1847 (1997).
46. Shirin, H. & Moss, S. F. Helicobacter pylori induced apoptosis. Gut 45, 592–594 (1998).
47. Gong, M., Ling, S. S. M., Lui, S. Y., Yeoh, K. G. & Ho, B. Helicobacter pylori γ-glutamyl transpeptidase is a pathogenic factor in the development of peptic ulcer disease. Gastroenterol. 139, 564–573 (2010).
48. Ricci, V., Giannouli, M., Romano, M. & Zarrilli, R. Helicobacter pylori gamma-glutamyl transpeptidase and its pathogenic role. World J. Gastroenterol. 20, 630–638 (2014).
49. Schmees, C. et al. Inhibition of T-cell proliferation by Helicobacter pylori γ-glutamyl transpeptidase. Gastroenterol. 132, 1820–1833 (2007). 50. Meister, A. & Anderson, M. E. Glutathione. Ann. Rev. Biochem. 52, 711–760 (1983).
51. Martensson, J., Jain, A. & Meister, A. Glutathione is required for intestinal function. Proc. Natl. Acad. Sci. 87, 1715–1719 (1990). 52. Circu, M. L. & Aw, T. Y. Glutathione and apoptosis. Free Radical Res. 42, 689–706 (2008).
53. Liu, C. et al. The effect of dietary glutathione and coenzyme Q10 on the prevention and treatment of inflammatory bowel disease in mice. Int. J. Vitam. Nutr. Res. 74, 74–85 (2004). 54. Liang, J. et al. Purification of Helicobacter suis strains from biphasic cultures by single colony isolation: influence on strain characteristics. Helicobacter 20, 206–216 (2015).
55. Baele, M. et al. Isolation and characterization of Helicobacter suis sp. nov. from pig stomachs. Int. J. Syst. Evol. Microbiol. 58, 1350–1358 (2008).
56. Joosten, M. et al. Diversity in bacterium-host interactions within the species Helicobacter heilmannii sensu stricto. Vet. Res. 44, 65 (2013). 57. Vermoote, M. et al. Antimicrobial susceptibility pattern of Helicobacter suis strains. Vet. Microbiol. 153, 339–342 (2011).
58. Flahou, B. et al. The local immune response of mice after Helicobacter suis infection: strain differences and distinction with Helicobacter pylori. Vet. Res. 43, 75 (2012).
59. Sugimoto, M., Ohno, T., Graham, D. Y. & Yamaoka, Y. Gastric mucosal interleukin-17 and -18 mRNA expression in Helicobacter pylori-induced Mongolian gerbils. Cancer Sci. 100, 2152–2159 (2009).
60. Crabtree, J. E. et al. Gastric mucosal cytokine and epithelial cell responses to Helicobacter pylori infection in Mongolian gerbils. J. Pathol. 202, 197–207 (2004).
61. Chirgwin, S. R. et al. Infection outcome and cytokine gene expression in Brugia pahangi-infected gerbils (Meriones unguiculatus) sensitized with Brucella abortus. Infect. Immun. 70, 5938–5945 (2002).
Acknowledgements
This work was supported by the Research Fund of Ghent University, Belgium, grant BOF08/GOA/004 and BOF14/GOA/010 and Fonds Wetenschappelijk Onderzoek Vlaanderen (grant FWO14/PDO/067). Technical assistance of Sofie De Bruyckere, Nathalie Van Rysselberghe and Sophie Callens is greatly acknowledged.
Author Contributions
F.H., R.D. and D.F. funding and study supervision. F.H., B.F., R.D. and D.F. study concept and design. E.D.B., B.F., M.J., G.Z. and M.S. acquisition of data. E.D.B., B.F. and D.F. analyses and interpretation of the data. E.D.B., B.F. and F.H. drafting of manuscript. R.D., M.J., G.Z., A.S. and F.P. critical revision of the manuscript.
Additional Information
Supplementary information accompanies this paper at http://www.nature.com/srep Competing financial interests: The authors declare no competing financial interests. How to cite this article: De Bruyne, E. et al. Oral glutathione supplementation drastically reduces Helicobacterinduced gastric pathologies. Sci. Rep. 6, 20169; doi: 10.1038/srep20169 (2016).
1Department of Pathology, Bacteriology and Avian Diseases, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium. 2Zoetis, Kalamazoo, MI 49007 USA. *These authors jointly supervised this work. Correspondence and requests for materials should be addressed to B.F. (email: bramflahou@hotmail. com)
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/